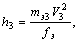 (1)
(1)
УДК 541.4:616.006.02
* 300000, Тула, ул.Менделеевская, 1, Научно-производственное предприятие Стек; тел./факс: (0872) 36-41-71;
** 300026, Тула, пр. Ленина, 108, НИИ новых медицинских технологий; лаборатории биофизики полей и излучений, медицинской генетики, медицинской СВЧ и КВЧ аппаратуры; тел./факс: (0872) 33-22-09;
E-mail: niinmt@mednet.com
*** 300026, Тула,
пр.Ленина, 125, Тульский государственный педагогический университет
им. Л.Н.Толстого, кафедра ботаники; тел.: (0872) 25-78-13
В связи с возникшим в последние годы пристальным научным интересом к продольным электромагнитным волнам (ПЭМВ) [1-3], которые предполагаются одним из основных носителей биоинформации в перцептивном канале [2, 3], перед физической, биологической и биофизической науками встал ряд вопросов концептуальной значимости, а именно: а) реализация методов и средств регистрации ПЭМВ; б) установление сходства или различия биологического воздействия на живые организмы ПЭМВ и обычных (поперечных) ЭМВ, причем сюда входит как аспект санкционированного лечебного воздействия, так и несанкционированного, сопутствующего негативного воздействия, вызывающего мутагенный эффект [4]; в) комплексные исследования лечебного (корригирующего) воздействия ЭМП, в том числе и полей ПЭМВ, на организм человека с позиций биокибернетического подхода [5] и т. п.
Из названного комплекса вопросов важнейшими, на наш взгляд, являются регистрация ПЭМВ и мутагенное воздействие на живой организм. Сложность регистрации ПЭМВ связана с их физической природой и поясняется ниже. Что же касается мутагенных эффектов при облучении биообъектов полями ПЭМВ, то этот момент рассматривается в следующих основных аспектах.
Во-первых, наиболее значима общая постановка вопроса, коль скоро речь идет о воздействии на организм на генетическом уровне полей ранее неисследованной физической природы, тем более, полагаемых основными составляющими в системе глобального биоинформационного обмена. Во-вторых, предполагая использование искусственно генерируемых ПЭМВ в терапевтических целях (одновре-менно помня и о возможности негативного санкционированного использования [1]), первостепенно необходимо исключить любую потенциальную возможность сопутствующих мутагенных эффектов. Наконец, существующие технические генераторы ПЭМВ [4] на сегодняшний день, а скорее всего и в ближней - до 2000-го года - перспективе, не позволяют получать строго продольные ЭМВ; в излучаемом ими спектре всегда присутствуют различные компоненты поперечных ЭМВ, в том числе и в КВЧ диапазоне, активно взаимодействующем с собственными ЭМП живых клеток [1]. Причем, если терапевтическое воздействие КВЧ ЭМП уже длительное время изучается и апробируется ([6, 7]; см. также специализированный журнал Миллиметровые волны в биологии и медицине, Москва), то планомерное исследование возможных мутагенных эффектов в этом диапазоне до сих пор не проводилось. Исключение последних из терапевтического диапазона - важнейшая задача высокочастотной физиотерапии.
Исходя из сказанного, авторы разработали принципиально
новый подход, названный ими методом биологического индикатора
(МБИ), позволяющий создать методологию регистрации ПЭМВ при одновременном
выявлении мутагенных эффектов.
Одна из сложностей регистрации ПЭМВ указана выше: это - на сегодняшний день - несовершенство схем генераторов продольных волн, а как следствие - излучение широкого спектра в виде композиции ПЭМВ и поперечных ЭМВ в диапазоне наибольшей активации собственных ЭМП клеток, то есть в КВЧ диапазоне.
Однако даже не эта причина является основной. Затруднена именно сама инструментальная возможность регистрации ПЭМВ, имея в виду обычно используемый в практике исследования полей набор радиофизических средств. Эту сложность наиболее просто представить, если исходить из той посылки, что в фундаментальном информационном коде Вселенной (понятие это в настоящее время является алгоритмически обоснованным и вошедшим в научный обиход информациологии, см., например, [1,8,9]) ПЭМВ априорно отведена роль носителя биоинформации на сверхнизкоинтенсивных энергетических уровнях.
Изучение ПЭМВ, связанное с именами Н.П.Хворостенко [10] и ряда других ученых [11], началось сравнительно недавно. Их существование не противоречит ни одному закону физики, более того, они удовлетворяют уравнениям Максвелла [1,10]. Однако для построения непротиворечивой теории ПЭМВ необходим не столько математический подход, сколько физический. В этом смысле позиции авторов укладываются в концепцию генерации ПЭМВ, исходя из представлений физического вакуума (ФВ), в настоящее время интенсивно развиваемую Тульской школой биофизики полей и излучений и биоинформатики [1-4, 9]. В данной концепции возникновение электрического поля интерпретируется как процесс поляризации ФВ, при котором часть сфероидообразных элементов ФВ (ЭФВ) занимает в пространстве положение (левое или правое), отличное от равновесного.
Исходя из основных типов осцилляций ЭФВ, в работе [3] определены базовые частотно-энергети-ческие характеристики ПЭМВ. Так, энергия основного уровня квантового осциллятора - условной вакуумной квазичастицы - равна
(1)где mэ3 - масса электрона для условий осцилляций со скоростью V3 продольной волны; fэ - частота соответствующих колебаний, а частота кванта продольного излучения составляет
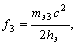 (2)
(2)где с - скорость света.
Из соотношений (1), (2) получаем, что h3 = 1,244301510-29 Джс 5,33105, где - постоянная Планка, а f3 1,821011 Гц. Отсюда следует, что: а) квант энергии ПЭМВ на пять порядков превосходит квант энергии поперечной ЭМВ; б) значение частоты f3, полученное для свободного электрона, указывает на область частот, в которой возможна передача информации между модами ЭМВ, а именно - ИК часть спектра, а по принципу симметрии и перманентности биополей - и КВЧ диапазон, - находящиеся в характерной частотной полосе дистанционного взаимодействия в живой природе.
Кроме того, из результатов [3] следует, что существуют специфические несущие частоты f3j для каждого j-го перцептивного канала информации на ПЭМВ. Резонансный характер взаимодействия поперечной и продольной мод ЭМВ на фиксированной частоте и его отсутствие на других частотах объясняет исчезновение спектра теплового шума в информационном (передаваемом) сигнале.
Таким образом, с одной стороны, в пользу особой
роли ПЭМВ как носителя биоинформации в структуре локальных и глобальных
информационных полей ноосферы [1] - говорит целый ряд факторов
(частотный диапазон, высокая энергетическая проникающая способность,
специфические несущие частоты для основных каналов, резонансный
характер взаимодействия и пр.), с другой - трудность регистрации
ПЭМВ радиофизическими приборами как раз объясняется тем, что ПЭМВ
распространяются малым числом высокоэнергетичных квантов. Поэтому
и возникает мысль о разработке методики не радиофизического, но
биологического индикатора ПЭМВ.
Из вышесказанного понятно, что МБИ предложен и используется в биофизике полей и излучений, что называется, не от хорошей жизни. Более того, сразу оговоримся, что МБИ не выдерживает никакой критики с точки зрения строгой методологии, но - на сегодняшний день - это есть единственный инструмент для решения задач, поставленных во введении. Суть же МБИ состоит в проведении параллельных экспериментов по облучению низших организмов неионизирующими квантами энергии поперечных ЭМВ диапазона КВЧ и ПЭМВ, широкий генерируемый спектр которых содержит и КВЧ составляющую. Регистрация ведется по факту устойчивого наблюдения мутагенного эффекта. Если данный эффект не наблюдается в серии опытов с облучением КВЧ ЭМВ, но устойчиво фиксируется при облучении электромагнитным полем от генератора ПЭМВ, то с определенной степенью доверительного риска можно говорить об индикации ПЭМВ по факту увеличения количества мутаций по сравнению с естественным уровнем.
В пользу принятия доверительного риска говорит инструментальная проверка генератора ПЭМВ с соответствующим квалификационным заключением ведущего академического института (г. Москва), свидетельствующая, что его высокочастотное электромагнитное излучение не является излучением поперечных ЭМВ. Ослабление же доверительности связано с широким (разрядным) спектром излучения генератора ПЭМВ, что приводит к стохастическим явлениям интермодуляции, разбросам поляризационных характеристик и пр.
И второй момент: метод МБИ опробован на Drosophila mеlanogaster. В то же время, как замечено в [4], поскольку системы репарации ДНК, а также системы образования и окисления промутагенов у насекомых и млекопитающих отличаются, то дальнейшим этапом в разработке МБИ явится проведение тестовых экспериментов на мутагенность на высших млекопитающих и на культуре клеток человека.
В проводимых комплексных исследованиях по МБИ, начатых в [4] и опирающихся на теорию и практику работы с ПЭМВ [1-3], в качестве наиболее удобного объекта для регистрации мутагенных факторов среды была выбрана Drosophila melanogaster (далее по тексту - дрозофила). К несомненным ее достоинствам относятся [4]: а) наличие значительного числа признаков, изменяющихся при мутации; б) способность активизировать мутагены, что важно для переноса использования методики на млекопитающих; в) возможность точно охарактеризовать и локализовать в геноме изменение под влиянием мутагена.
Для практической реализации МБИ используется известное свойство [4] дрозофил, гетерозиготных по одному или более рецессивным генам: под влиянием различных внешних раздражителей, в частности ионизирующего излучения, у последних появляются мозаичные пятна на участках тканей, соответствующих действию этих генов, - на общем фоне дикого фенотипа. Считается, что основным механизмом возникновения пятен является соматический кроссинговер; также заметна роль элиминации хромосом, точковых мутаций, аббераций хромосом и морфозов [4]. Таким образом, наиболее эффективным методом анализа влияния излучений на мутагенность является исследование их способности вызывать у дрозофилы соматический мозаицизм, а при объективировании на рецессивные гены удобство исследования связано с тем, что для них гомозиготный эффект фиксируется на относительно малых участках тела.
Еще раз подчеркнем, что используются неионизирующие излучения нетепловой (биоинформацион-ной) интенсивности с плотностью мощности порядка 1...10 мВт/см2; при этих значениях нагрев биообъекта не превышает 0,1оС, то есть не является физиологически значимым.
Сущность соматического кроссинговера состоит в том,
что он осуществляется при митотическом делении клеток соматических,
преимущественно эмбриональных тканей. У дрозофил соматический
кроссинговер фиксируется следующим образом. В эксперименте участвуют
самки, гетерозиготные по рецессивным генам у (желтая окраска
щетинок) и sn (опаленные щетинки): 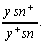
В случае, если в соматических клетках в профазе происходит перекрест в данной паре хромосом, на теле могут появляться пятна с рецессивными признаками. При этом в зависимости от того, где произойдет перекрест между геном sn и центромерой или между генами y и sn, появятся либо двойные пятна (sn и y), либо одиночные (только sn или y), в которых проявляется действие этих генов, оказывающихся в гомозиготном состоянии.
Процесс соматического кроссинговера у дрозофил хорошо изучен. Отметим, что он возможен не только для половых хромосом, но и для аутосом, осуществляясь как у самок, так и у самцов. Изучение соматического кроссинговера у дрозофилы со сцепленными Х-хромосомами позволило сделать вывод, что кроссинговер осуществляется на стадии четырех хроматид.
Существующие концепции, объясняющие механизмы взаимодействия
продольных и поперечных ЭМВ с клеточными и субклеточными структурами, -
биофизическая и биохимическая основа соматического кроссинговера
- изложены в [1-7]. Другое дело, что исследование этих механизмов,
теоретическая база и эксперимент находятся в начальной стадии
исследования.
Экспериментальное исследование воздействия на соматический кроссинговер дрозофилы излучения ПЭМВ было проведено авторами в [4]. Исследование выполнялось на гетерозиготных дрозофилах по двум Х-сцепленным рецессивным генам: y(yellow) и sn3(singed). Эти гены находятся в трансположении; вследствие гетерозиготности они фенотипически не проявляются. При структурных нарушениях в хромосоме гены приходят в гомозиготное состояние и становятся видимыми по форме и цвету щетинок (см. выше). В зависимости от места разрыва хромосомы возникают клетки разных генотипов, то есть проявляются пятна разных типов. Разрыв между геном sn и центромерой приводит к образованию двойного пятна: две соседние макрохеты проявляют действие генов: одна - yellow, другая - singed. Разрыв между генами sn и y, а также двойной кроссинговер приводит к появлению одиночных пятен yellow и singed соответственно. При такой схеме скрещивания можно судить о мутагенности излучения ПЭМВ по повышению частоты соматического мозаицизма [4]:
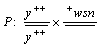
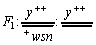
В работе [4] приведено описание эксперимента по воздействию на дрозофилы в стадии личинок излучением генераторов ПЭМВ, созданных в лаборатории биофизики полей и излучений НИИ новых медицинских технологий МЗ РФ (г. Тула), а также статистически обработанные результаты тестирования генераторов ПЭМВ на индуцирование соматических мутаций в системе генов y и sn3 у гетерозиготных самок дрозофил.
Заметные результаты были обнаружены при облучении ПЭМВ в варианте генерации Е-10. Процент мозаичных пятен превышал контроль в 2,5 раза. Математическая обработка по методике [12] показала, что достоверная оценка соматического мозаицизма в варианте Е-10 показывает безусловный мутагенный эффект (U-критерий равен 2,20 >1,96). При облучении четко проявлялись различные типы морфозов, численно незначительно превышавшие фоновые показатели. Наблюдалась гибель порядка 25% личинок после облучения.
Следует отметить по результатам экспериментов в [4], что наличие мутагенного эффекта в значительной степени зависит от режима генерации (частоты, модуляции и пр.) - типа генератора ПЭМВ.
Параллельно с облучением дрозофил продольными волнами [4] проводился контрольный эксперимент по облучению поперечными ЭМВ. Прочие условия эксперимента были адекватными. Виргинных самок генотипа y+/y+ (yellow, 1-0; 0; рецессивный ген в Х-хромосоме, обуславливающий развитие желтой окраски тела и щетинок) скрещивали массово с самцами wsn/y (white, 1-1,5; белая окраска глаз; sn-singed,1-21,0; щетинки извитые; оба гена - также рецессивные, Х-хромосомные).
В каждую пробирку 20 мм, содержащую 10 мл стандартной питательной среды, помещались по 5 самок и 3 самца. Через каждые два дня родители пересаживались на свежую питательную среду, а среда, содержащая яйца и одно-двухдневные личинки (первая стадия), подвергались облучению.
Облучение ЭМВ (стандартный генератор ГЧ-115) производилось на расстоянии пробирок от корпуса генератора 15 см в течение 30 минут. Эксперимент содержал 6 этапов (опытов), проводившихся при постоянной мощности излучения; частота модуляции имела следующие дискретные значения: 1 = 37,5 ГГц; 2 = 37,0 ГГц; 3 = 36,5 ГГц; 4 = 36,0 ГГц; 5 = 35,5 ГГц; 6 = 35,0 ГГц. Температура в помещении выдерживалась в пределах 20-20,5оС, а атмосферное давление - 94-100 кПа. Эксперимент проводился в течение 3 месяцев. После облучения партии пробирок помещались в термостат с температурой 230,5оС.
Вылет дрозофил начинался к восьмому дню; по мере
вылета особей просматривали под микроскопом МБС-9 в падающем свете.
У гетерозиготных самок 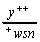 регистрировались
мутантные yellow либо singed макрохеты на голове
и тораксе.
регистрировались
мутантные yellow либо singed макрохеты на голове
и тораксе.
По изложенной выше методике в ходе эксперимента проводился анализ влияния ЭМВ указанных частот на ДНК модельного организма Drosophila melanogaster - способности их к мутагенному эффекту. Полученные данные представлены в табл. 1.
В соответствии с данными эксперимента (табл. 2)
можно достоверно считать, что облучение КВЧ ЭМП в диапазоне частот
от 35 до 37,5 ГГц не индуцирует мутаций в системе генов y
и sn (U1 = 0,220 < 1,96;
U2 = 0,637 < 1,96; U3 =1,54 < 1,96;
U4 = 0,556 <1,96; U5 = 0,876 <1,96;
U6 =1,037 < 1,96). При облучении
проявляются различные типы морфозов, число которых не превышает
фоновые показатели.
Исходя из теоретических проработок и сравнения экспериментальных данных, полученных в настоящей и предыдущей [4] работах, можно сделать следующие выводы.
1. Экспериментально подтверждено существование продольных ЭМВ, наличие которых ранее предполагалось алгоритмически [1-3], причем подтверждается теоретический вывод о том, что ПЭМВ низкой интенсивности практически не взаимодействуют с материальными средами (объектами) неживой природы, но воздействуют на геном живого организма, что отвечает концепции [1] об особой роли ПЭМВ в биоинформационном обмене.
2. Поперечные ЭМВ в диапазоне 35-37,5 ГГц не вызывают мутагенного эффекта у Drosophila melanogaster, о чем свидетельствует достоверная оценка соматического мозаицизма в эксперименте. Отсутствие мутаций на дискретных частотах в диапазоне от 2 до 8 мм при КВЧ излучении низкой интенсивности подтверждается практикой использования КВЧ терапевтических аппаратов [6, 7]. Наконец, практика работы с КВЧ радиолокационными станциями, в основном специального назначения (системы наведения и сопровождения), работающими на различных частотах КВЧ диапазона, также подтверждает отсутствие мутаций (послед-ствия от облучения КВЧ ЭМП и СВЧ ЭМП высокой мощности имеют иную природу). Однако в целях чистоты экспериментов, а главное - безопасности КВЧ терапии на нетрадиционных частотах, желательной является крупномасштабная проверка на мутагенность всего КВЧ диапазона: 30-300 ГГц.
3. Поскольку системы репарации ДНК, а также системы образования и окисления промутагенов у насекомых и млекопитающих отличаются, то продолжение цикла исследований, начатого в [4] и настоящей работе, целесообразно провести в виде тестирования на мутагенность высших млекопитающих и культуры клеток человека.
4. При всей своей неадекватности и отсутствии строго методологического подхода метод биологического индикатора является на сегодняшний день единственным практическим способом регистрации физических полей низкой интенсивности, фиксация которых по той или иной причине затруднена инструментальными методами, в частности стандартной радиофизической аппаратурой.
Таблица 1
Примечание: результаты математической обработки данных
эксперимента по методике [12] приведены в табл 2
Таблица 2
облучения ,ГГц |
 |
||||
Примечание: Р = процент мозаичных пятен; - функция
распределения Фишера;
Авторы выражают благодарность сотрудникам Тульского государственного
педагогического университета им. Л.Н. Толстого В.В. Воронову
и Р.А. Сидорову за помощь в проведении экспериментов.
 - доверительный
интервал (U = 1,96); n - число просмотренных
особей (no - в опыте, nk -
в контроле); S - стандартная ошибка; Sk-o
- cтандартная ошибка разности между k (контроль)
и o (опыт), U - критерий вероятности,
- доверительный
интервал (U = 1,96); n - число просмотренных
особей (no - в опыте, nk -
в контроле); S - стандартная ошибка; Sk-o
- cтандартная ошибка разности между k (контроль)
и o (опыт), U - критерий вероятности,

1. Взаимодействие физических полей с живым веществом / Е.И.Нефедов, А.А.Протопопов, А.Н.Семенцов, А.А.Яшин; Под ред. А.А.Хадарцева.- Тула: Изд-во Тульск. гос. ун-та, 1995.- 180 с.
2. Афромеев В.И., Нефедов Е.И., Протопопов А.А., Хадарцев А.А., Яшин А.А. Параметры информационного канала на продольных электромагнитных волнах // Вестник новых медицинских технологий.- 1996.- Т. III, ¦ 4.- C. 31-32.
3. Нефедов Е.И., Протопопов А.А., Семенцов А.Н., Федорищев И.А., Яшин А.А. Параметры перцептивного канала информации на продольных электромагнитных волнах // Вестник новых медицинских технологий.- 1995.- Т. II, ¦ 1-2.- C. 21-27.
4. Богданов В.П., Воронов В.В., Сидоров Р.А., Яшин А.А. Исследование методом соматической рекомбинации дрозофил, подвергшихся воздействию продольных электромагнитных волн // Вестник новых медицинских технологий.- 1995.- Т. II, ¦ 3-4.- C. 6-9.
5. Афромеев В.И. Соотношение биологического, физического и математического в реализации лечебно-диагностического воздействия высокочастотных полей // Вестник медицинских технологий.- 1997.- Т.IV, ¦ 1-2.- C. 16-22.
6. Миллиметровые волны в медицине: Сб.ст., Т. 1, Т.2 / Под ред. Н.Д.Девяткова и О.В.Бецкого.- М.: Изд-во Ин-та радиотехн. и электрон. РАН, 1991.- 585 с.
7. Миллиметровые волны в биологии и медицине (библиография).- М.: Изд-во ЗАО МТА-КВЧ, 1996.- 39 с.
8. Герловин И.Л. Основы единой теории всех взаимодействий в веществе.- Л.: Энергоатомиздат, 1990.- 432 с.
9. Нефедов Е.И., Яшин А.А. Информационная безопасность человека как категория ноосферной экологии // Вестник новых медицинских технологий.- 1995.- Т. II, ¦ 3-4.- C. 37-45.
10. Хворостенко Н.П. Продольные электромагнитные волны // Изв. ВУЗов. Сер. Физика.- 1992.- ¦ 3.- C. 24-29.
11. Герценштейн М.Е., Болошин И.А. О возможном физическом носителе перцептивного канала информации // Вопросы магнитостатики и электродинамики.- М., 1978.- С. 70-75.
12. Методические рекомендации по применению
соматического мутагенеза у Drosophila melanogaster в качестве
тест-системы для ускоренного определения канцерогенов.- М.: МЗ
СССР, 1982.
The authors suggest the method called biological
detector (MBD), based on gene responsivity to physical low-intensity
fields which can't be recorded with routine technical means. As
longitudinal electromagnetic waves (LEMWS) play a specific role
in local and global bioinformation exchange, it has been studied
an effect of LEMW-radiation on somatic crossing-over of Drosophila
melanogaster, the radiation being of two types (i.e., radiation
generated with a tested generator and typical LEMW one in an EHF
range). The findings show that LEMWs exist and induce a mutational
effect, but electromagnetic EHF waves do not induce such an effect.