УДК 681.51:621.391.008.05
КОРРЕЛЯЦИОННЫЙ ПОДХОД И РОЛЬ ФИЗИОЛОГИЧЕСКИХ
РИТМОВ В ОБЪЯСНЕНИИ ЭФФЕКТОВ ВЗАИМОДЕЙСТВИЯ ЭЛЕКТРОМАГНИТНЫХ ПОЛЕЙ
С ЖИВЫМ ОРГАНИЗМОМ
В. И. АФРОМЕЕВ*, Т. И. СУББОТИНА**,
А. А. ЯШИН**
* 300000, Тула, ул.
Менделеевская, 1, Научно-производственное предприятие Стек; тел.:
(0872) 36-41-71
** 300026, Тула, пр. Ленина,
108, НИИ новых медицинских технологий, тел./факс: (0872) 33-22-09;
Е-mail: root@niinmt.mednet.com
1. Введение
Вопрос о механизме активации собственных электромагнитных
полей (ЭМП) клеток организма при воздействии крайневысокочастотных
(КВЧ) внешних полей (санкционированных или несанкционированных)
является на сегодняшний день наиболее важным и наименее изученным
в контексте КВЧ терапии, а также медико-биологического эксперимента
на животных. Причина сложившейся ситуации видится троякой: а)
достаточно малый - по времени и программной реализации - период
исследований по данной тематике; у нас в стране более или менее
серьезные исследования начались лишь во второй половине 80-х гг.,
а за рубежом к ним только приступают; б) преобладание радиофизического
подхода, в то время как здесь более адекватен подход биофизический;
причиной этого является определенный субъективизм основной группы
исследователей, ранее занимавшихся исключительно вопросами радиофизики
и технической электродинамики; в) самое основное связано с тем,
что величина излучения клетки на частотах длинноволновой части
КВЧ диапазона (порядка 40 ГГц) составляет Ркл 10-23 Вт,
причем накладываясь друг на друга, поля ансамбля клеток дают общее
ЭМП со спектром шумового характера. Это предъявляет требования
к чувствительности измерительной аппаратуры, превосходящие аналогичные
для современных радиотелескопов.
Сказанное стимулирует поиски гипотетических механизмов
активации собственных ЭМП клеток.
2. Анализ возможных объяснений механизма
взаимодействия собственных полей клеток с внешними полями
Несмотря на многочисленные публикации по рассматриваемому
вопросу, отсутствует анализ основных объяснений механизма активации.
Попробуем восполнить этот пробел. Одной из первых и пользующихся
наибольшей известностью является гипотеза о когерентном
характере активации (Биофизическая школа акад. Н.Д. Девяткова
- Институт радиотехники и электроники РАН: М.Б. Голант, О.В. Бецкий
и др.) [1]. Согласно этой гипотезе, микроструктура клеточных мембран
(совокупность мембран митохондрий) обеспечивает возникновение
дипольной компоненты клетки. Такой колеблющийся электрический
диполь 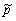 = ро sin t,
связанный с акустическим колебанием мембраны, является клеточным
генератором собственного КВЧ ЭМП клетки. В здоровой клетке характер
колебаний является стохастическим (особенно с учетом взаимодействия
ЭМП ансамбля клеток), а спектральная характеристика поля близка
к шумовой с некоторой средней интенсивностью
= ро sin t,
связанный с акустическим колебанием мембраны, является клеточным
генератором собственного КВЧ ЭМП клетки. В здоровой клетке характер
колебаний является стохастическим (особенно с учетом взаимодействия
ЭМП ансамбля клеток), а спектральная характеристика поля близка
к шумовой с некоторой средней интенсивностью 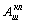 (рис. 1). При патологических изменениях в клетке реакция
последней выражается в возрастании интенсивности генерации по
отношению к
(рис. 1). При патологических изменениях в клетке реакция
последней выражается в возрастании интенсивности генерации по
отношению к 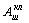 в относительно узких полосах
спектра, например, как показано на рис. 1, на участках 2 fp1
или 2 fp2, с некоторыми центральными
частотами fp1 или fp2.
Можно предположить, что, с учетом той или иной формы нарушения
метаболических процессов в клетке, возрастание интенсивности генерации
в относительно узких полосах
спектра, например, как показано на рис. 1, на участках 2 fp1
или 2 fp2, с некоторыми центральными
частотами fp1 или fp2.
Можно предположить, что, с учетом той или иной формы нарушения
метаболических процессов в клетке, возрастание интенсивности генерации
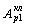 или
или 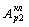 связано
с перераспределением свободной энергии и притоком ее на
участки спектра возмущения (простейший и понятный аналог на макроорганизменном
уровне - повышение температуры тела при воспалительном заболевании).
связано
с перераспределением свободной энергии и притоком ее на
участки спектра возмущения (простейший и понятный аналог на макроорганизменном
уровне - повышение температуры тела при воспалительном заболевании).
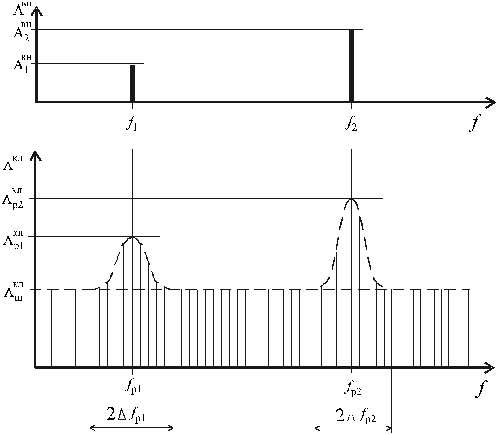
Рис.1. К гипотезе
о когерентном характере активации собственных ЭМП клеток
Если теперь наложить на собственное ЭМП клетки когерентное
внешнее ЭМП с частотами f1 или f2
(на рис. 1 для упрощения показаны монохроматические колебания),
то поле клетки входит с внешним полем в резонанс, что, по принципу
цепной реакции, характерной для биофизических и биохимических
процессов в клетках, увеличивает приток свободной энергии, а значит
и интенсивность колебаний 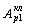 или
или 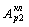 резонансно возрастает, биохимические процессы в клетке интенсифицируются,
что приводит к выздоровлению клетки.
резонансно возрастает, биохимические процессы в клетке интенсифицируются,
что приводит к выздоровлению клетки.
Согласно гипотезе когерентного резонанса
собственно процесс является биоинформационным, поскольку
для осуществления цепной реакции в клетке интенсивность Авн
внешнего ЭМП особой роли не играет; важно, в основном, его информационное
содержание (частота, модуляция, поляризация и т. п.).
Слабым местом этой концепции является отсутствие
сколь-либо аргументированного выбора дискретных частот внешнего
ЭМП, используемого в широкой терапевтической практике: 3-4 терапев-тические
монохроматические частоты в диапазоне от 2 до 8 мм. Возможный
(сугубо субъективный) вариант выбора этих частот рассмотрен в
[2].
Второй, неизученный даже в нулевом (гипо-тетическом)
приближении момент - связь частот возмущения генерации клеток
с нозологией заболевания. И, наконец, нет никакой объяснимой связи
между эффектами лечебного воздействия на больной организм, корректирующего
(поддерживаю-щего) воздействия на здоровый организм и все это
вместе - с выбором четко дискретных, количественно ограниченных
терапевтических частот.
В еще большей степени биоинформационный характер
активизации клеточных ЭМП утверждается в подходе Тульской школы
биофизики полей и излучений и биоинформатики - НИИ новых медицинских
технологий МЗ РФ (В.П. Богданов, Е.И. Нефе-дов, А.А. Протопопов,
А.А. Хадарцев, А.А. Яшин и др.) [3]. Концепция последней
в настоящее время формируется в русле общей концепции теории единого
информационного поля ноосферы (Е.И. Нефе-дов, А.А. Яшин)
и особой роли продольных электромагнитных волн (ПЭМВ) в биоинформационном
обмене. Многое здесь зависит от результатов окончания первого
этапа теоретических и экспериментальных исследований (Исследования
проводятся с 1995 года в рамках ряда НИОКР совместно с ИРЭ РАН
и другими организациями).
Можно отметить в данном вопросе активно проводимые
исследования в Институте биофизики клетки РАН (г. Пущино-на-Оке;
руководитель работ Н.К. Чемерис). В результате теоретических
и экспериментальных исследований (см., например, [4]) представителями
Пущинской биофизической школы сформирована концепция стохастического
резонанса с особой ролью двойного резонанса на частотах КВЧ
несущей и низкочастотного модулирующего сигнала; последний, по
всей видимости, находится в частотной области основных физиологических
ритмов организма.
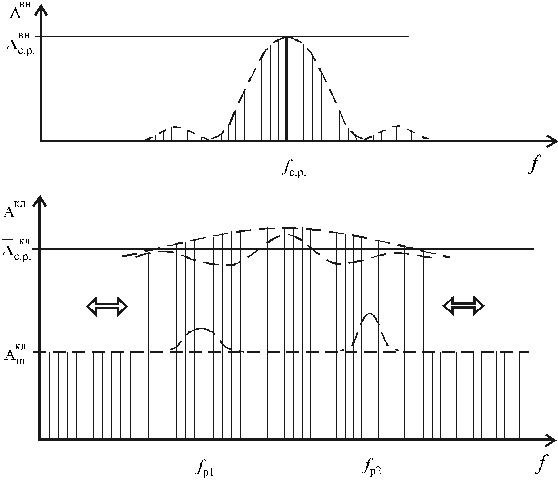
Рис. 2. К концепции
стохастического резонанса в
активации собственных ЭМП клеток
В первом приближении этот процесс графически можно,
очевидно, представить (рис. 2). Здесь, как мы видим, локального
возрастания интенсивности генерации (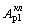 или
или 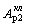 ) может и не быть, но воздействие
внешнего ЭМП в форме модулированного сигнала с амплитудой
) может и не быть, но воздействие
внешнего ЭМП в форме модулированного сигнала с амплитудой 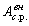 и с некоторой средней частотой f c.p. приводит
к возрастанию амплитуды стохастических колебаний клетки
и с некоторой средней частотой f c.p. приводит
к возрастанию амплитуды стохастических колебаний клетки 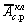 по отношению к уровню
по отношению к уровню 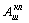 в достаточно широкой
полосе. То есть перераспределение свободной энергии метаболизма
клетки происходит не в виде частотного резонанса, но резонанса
энергетического в достаточно широком частотном спектре. Поэтому
один из предварительных, само собой напрашивающихся выводов из
концепции стохастического резонанса говорит не о биоинформационном,
но биоэнергетическом характере активации генерации клеток
внешним ЭМП. В то же время из эксперимента [4] однозначно следует,
что энергетичность внешнего электромагнитного воздействия роли
не играет; достаточно лишь относительно низкого (порогового) уровня.
в достаточно широкой
полосе. То есть перераспределение свободной энергии метаболизма
клетки происходит не в виде частотного резонанса, но резонанса
энергетического в достаточно широком частотном спектре. Поэтому
один из предварительных, само собой напрашивающихся выводов из
концепции стохастического резонанса говорит не о биоинформационном,
но биоэнергетическом характере активации генерации клеток
внешним ЭМП. В то же время из эксперимента [4] однозначно следует,
что энергетичность внешнего электромагнитного воздействия роли
не играет; достаточно лишь относительно низкого (порогового) уровня.
В принципе это вполне согласуется с биологическим
механизмом цепной реакции образования свободной энергии метаболизма
клетки, однако для отказа от информационной компоненты необходимо
провести большую серию экспериментов по всем диапазонам ЭМП, играющим
жизненно важную роль в функционировании клеток. Наиболее привлекательной
стороной данной концепции является выраженный экспериментальный
подход.
Известен ряд других подходов, например, развиваемый
И. В. Родштатом (Фрязино, ИРЭ РАН) [5-7] (См.
также публикации в Вестнике новых медицинских технологий за 1994-97
гг.). Зарубежные исследования пока не носят
фундаментального характера, но привлекают хорошую экспериментальную
базу.
3. Корреляционный подход к объяснению
механизма активации собственных полей
клеток
При подходе к исследованию механизма активации мы
исходим, прежде всего, из информационной основы биофизикохимических
процессов жизнедеятельности [3] и всеобщего биокибернетического
подхода к организации сложных (само-организующихся, нелинейных,
синергетических и пр.) систем [8].
Как видно из изложенного выше, основные противоречия
системного характера возникают в предпочтении тех или иных
сочетаний процессов, определяемых ключевыми понятиями: биофизический
процесс, биохимический процесс, свободная энергия метаболизма
клетки, биоинформационный и/или биоэнергетический характер воздействия
внешнего ЭМП, собственное ЭМП клетки, акустические колебания и
пр.
Очевидно, что не имеет смысла спорить о первенстве
(или доминанте) биохимического или биофизического процессов в
создании собственного ЭМП клетки. Понятно, что свободная энергия,
необходимая клетке в том числе и для генерации КВЧ ЭМП, вырабатывается
при биологическом окислении в митохондриях. Важнейшую роль
здесь играет аденозинтрифосфат (АТФ). А далее эта энергия
передается в клетку химическим путем. Даже не важно, что считать
базовым носителем этой энергии: электроны, как то считает классическая
клеточная термодинамика [9], или протоны в представлении новых
концепций биоэнергетики клетки [10]. Главное здесь то, что биохимические
и биофизические процессы вообще неразрывно связаны в функционировании
клетки, коль скоро речь идет о превращениях энергии.
Подойдем с другой стороны. Считается доказанным,
что клеточные диполи генерируют именно КВЧ ЭМП, причем здесь значима
роль акустоэлектронных процессов колебаний; в [3] приведена
сводка расчетных результатов, где фигурируют и мощности клеточных
полей, и частоты, попадающие в миллиметровую часть КВЧ диапазона
и пр. Но почему клетка генерирует именно в этом, техническом,
но совсем не характерном для живой природы диапазоне? Все живое
на Земле cвоим возникновением, эволюцией, существованием и тенденциями
развития обязано лучистой энергии Солнца; опосредованно - и донные
рыбы, и бактерии глубинных земных пластов. Живые же организмы,
освещаемые солнечным светом весь день, - тем более. Поэтому
все биохимические и биофизические процессы такого организма синхронны,
настроены на контроль по ЭМП спектра солнечных лучей, отфильтрованных
атмосферой Земли, в котором практически отсутствует КВЧ компонент.
А если перенестись в те далекие биогеохимические эпохи (основная
терминологическая единица в учении В.И. Вернадского), когда
зарождалась на Земле жизнь, формировалась клетка, почти неизменной
доставшаяся нам, то сквозь перенасыщенный пар безозоновой атмосферы
до поверхности доходили не все составляющие спектра видимого света.
Таким образом, все клеточные и субклеточные структуры настроены
на УФ, отчасти - ИК диапазоны [3], а их сродство к КВЧ ЭМП в контексте
(к определяющим основные процессы жизнедеятельности) взаимодействия
с внешними ЭМП является логически неувязанным.
Опять же в существующих концепциях недостаточно
ясно увязана полевая специфика клетки со сложностью состава окислительной
дегидрогеназы (свыше 200 ферментов) и двойственностью самого окислительно
процесса в митохондриях: ферментативного и неферментативного свободнорадикального.
И практически не исследована роль матриксов - жидкой матричной
среды существования клеточных структур и субструктур: от межклеточной
среды до внутримитохондриального матрикса.
Сказанное выше, с учетом теоретически адекватных
и экспериментально подтвержденных факторов, позволяет построить
гипотетическую корреляционную модель активации собственных
ЭМП клеток организма, суть которой сводится к следующему.
Естественным биологическим полем живого организма
является ЭМП диапазонов от ИК до УФ, а возможно и более коротковолновое.
В данном случае мы отвлекаемся от механизма его генерации; возможные
варианты последней проанализированы в [3]; может быть - согласно
гипотезе Г.Н. Пет-раковича [10] - это поле изначально
связано с цитохромными процессами колебания электронов в системе
четырех связанных между собою атомов железа:
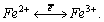
Главное, что колебания S1(t)
генерируются (рис. 3), а само их наличие необходимо для реализации
биокибернетического принципа обратной связи организма со средой
[8].
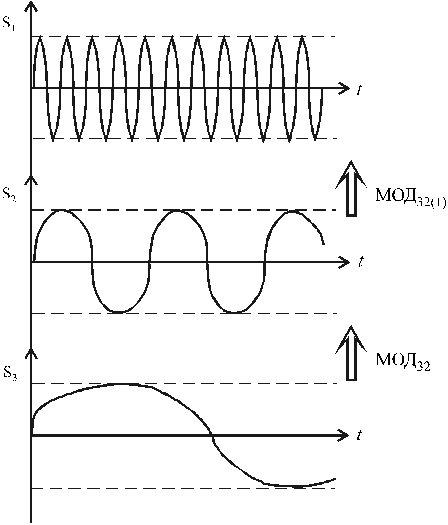
Рис. 3. К принципу
работы корреляционной модели в активации собственных ЭМП клеток
Вторым и тоже естественным полем организма являются
поля, обусловленные физиологическими ритмами S3(t);
это низкочастотные колебания от долей герца до сотен (тысяч) герц,
физически реализуемые как акустоэлектрические колебания. И это
все, другие жизненно важные для жизнедеятельности организма поля
можно не привлекать. Но здесь возникает, с точки зрения радиофизической,
весьма существенный вопрос: в пределах организма как (повторимся)
системы биокибернетической и биофизической одновременно все поля
должны быть связаны; в данном случае эта корреляционная связь
единственно представляется как модуляция (в модели - амплитудная,
но в действительности носящая очень сложный, комбинированный характер).
Однако непосредственная модуляция МОД31(S3/S1)
вряд ли возможна, ибо слишком иерархически удалены друг от друга
соответствующие биоструктуры - среды генерации и распределения
ЭМП: колебания S1(t) существуют в микроструктурах
клеток; на уровне, например, ферропротеидов, а средой распространения
акустоэлектрических колебаний S3 является, преимущественно,
структура (решетка) межклеточной среды.
Поэтому (и только для этого, но не для отклика на
воздействие внешних, природных ЭМП) в организме существуют колебания
S2(t) - скорее всего как колебания заряженных
клеточных мембран, часть спектра которых, скорее всего - длинноволновая,
совпадает с КВЧ диапазоном. То есть их назначение в естественном
биофизическом процессе - корреляционно-связующее, медианно-модулирующее.
Соответственно двойная модуляция МОД32 (S3/S2)
МОД32(1) (S3/S2/S1)
приводит к расширению спектра итогового сигнала А32(1)
(f).
Изложенная концепция является непротиворечивой,
доказательство последнего требует отдельного и аргументированного
рассмотрения.
4. Заключение
В общих чертах пока можно утверждать, что
непротиворечивость изложенной гипотезы соотносится с основными
выводами из существующих концепций. Так биоинформационная сущность
активации или, что то же самое - низкоинтенсивная биоэнергетическая
основа ее, проявляется, с одной стороны, в попадании в область
некоторых характерных частот спектра А32(f)
с несущей 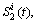 с другой - наложением
на квазишумовой спектр А32(1) (f) или
А32 (f). Явление двойного резонанса при
воздействии внешним КВЧ сигналом, модулированным низкочастотным
сигналом, в концепции стохастического резонанса также является
легко объяснимым. Цепная реакция биохимических процессов, инициируемая
внешним ЭМП, приводящая к интенсификации выделения свободной энергии
и пр., не противоречит основным доводам. Перечисление можно продолжить.
с другой - наложением
на квазишумовой спектр А32(1) (f) или
А32 (f). Явление двойного резонанса при
воздействии внешним КВЧ сигналом, модулированным низкочастотным
сигналом, в концепции стохастического резонанса также является
легко объяснимым. Цепная реакция биохимических процессов, инициируемая
внешним ЭМП, приводящая к интенсификации выделения свободной энергии
и пр., не противоречит основным доводам. Перечисление можно продолжить.
Однако основной вывод видится в констатации своего
рода служебной роли клеточных ЭМП диапазонов, адекватных КВЧ диапазону.
Как то мы часто наблюдаем в нашем многогранном мире, природа,
создавая свои творения, не всегда может предположить, что рукотворное
детище созданного ею Homo sapiens может вмешиваться в самоорганизующуюся
систему организма. Отсюда, кстати, и все чаще поднимаемый вопрос
о ноосферной экологии [11,12].
Литература
1. Девятков Н.Д., Голант М.Б., Бецкий О.В.
Гипотеза о взаимосвязанности воздействий когерентных волн малой
мощности КВЧ, ИК, оптического и УФ диапазонов на функционирование
клеток // В кн.: Миллиметровые волны в медицине: Сб.ст. Т. 2
/ Под ред. Н.Д. Девяткова и О.В. Бец-кова.- М.: Изд-во
Ин-та радиотехн. и электрон. РАН, 1991.- С. 349-362.
2. Хадарцев А.А., Яшин А.А. Новые медицинские
технологии лечения заболеваний внутренних органов и их аппаратурное
обеспечение // Вестник новых медицинских технологий.- 1996.- Т. III,
¦ 2.- С. 6-9.
3. Взаимодействие физических полей с живым
веществом / Е.И. Нефедов, А.А. Протопопов, А.Н. Семенцов,
А.А. Яшин; Под ред. А.А. Хадарцева.- Тула: Изд-во Тульск.
гос. ун-та, 1995.- 180 с.
4. Ганеев А.Б. Особенности действия модулированного
электромагнитного излучения крайневысоких частот на клетки животных:
Автореф.дисс. ... канд.физ.-мат.наук.- Пущино: Ин-т теорет. и
эксперимент. биофизики РАН, 1997.- 21 с.
5. Родштат И.В. КВЧ-модуляция процессов функционирования
и смерти корпоральных и мозговых тканей / Препринт ¦ 8(596)
ИРЭ РАН.- М.: Изд-во Ин-та радиотехн. и электрон. РАН, 1994.-
36 с.
6. Родштат И.В. Некоторые вопросы терминальных
состояний, процесса смерти, субстанций человека в контексте КВЧ-терапии
/ Препринт ¦ 3(591) ИРЭ РАН.- М.: Изд-во Ин-та радиотехн.
и электрон. РАН, 1994.- 23 с.
7. Родштат И.В. Вопросы объективизации некоторых
дискуссионных психофизиологических феноменов, предположительно
модулируемых КВЧ-воздействием / Препринт ¦ 2(602) ИРЭ РАН.-
М.: Изд-во Ин-та радиотехн. и электрон. РАН, 1995.- 23 с.
8. Афромеев В.И. Соотношение биологического,
физического и математического в реализации лечебно-диагностического
воздействия высокочастотных полей // Вестник новых медицинских
технологий.- 1997.- Т. IV, ¦ 1.- C. 18-25.
9. Вестерхофф Х., ван Дам К. Термодинамика
и регуляция превращений свободной энергии в биосистемах: Пер.с
англ.- М.: Мир, 1992.- 686 с.
10. Петракович Г.Н. Ядерные реакции в живой
клетке // Русская Мысль.- 1993.- ¦ 3-12.- С. 66-73.
11. Нефедов Е.И., Яшин А.А. Информационная
без-опасность человека как категория ноосферной экологии // Вестник
новых медицинских технологий.- 1995.- Т. II, ¦ 3-4.- С. 51-62.
12. Нефедов Е.И., Яшин А.А. Электромагнитная
основа в концепции единого информационного поля ноосферы // Философские
исследования.- 1997.- ¦ 1.- C. 5-74.
ABOUT A POSSIBLE CORRELATIVE MECHANISM OF
AN ACTIVATION OF PROPER ELECTROMAGNETIC FIELDS IN ORGANISM
CELLS UNDER AN EXTERNAL EXPOSURE TO RADIATION
V.I.AFROMEEV, T.I.SUBBOTINA, A.A.YASHIN
Summary
An activation mechanism of proper EMFs of cells
under external EHF fields is discussed. It is shown that main
concepts existing on the above issue can be reduced, from one
hand, to coherent resonance or guasienergy resonance and, from
the other hand, to a bioinformation or bioenergy dominant. The
authors suggest a correlative mechanism using 3-level model of
internal EMF in an organism, which intercorrelate according to
modularity. EHF EM fields link UV and low-frequency acoustoelectric
fields that represent main physiological rhythms of the organism
in a single but hierarchically co-ordinating structure of the
object. The concept needs subsequent development.