УДК 612.822.3
В середине 80-х годов в рамках изучения оперантивно-обусловленных форм вызванной нейронной активности был установлен ряд фактов, свидетельствующих о существовании особого механизма воздействия биологически активных веществ на клетки головного мозга [1, 2]. Выяснилось, что некоторые органические соединения, не оказывающие прямого влияния на реализацию специализированной функции нервной клетки, приобретают такого рода способность в условиях биоуправляемого микроионофореза (Lself-application approach¦).
Первоначально эта довольно странная закономерность рассматривалась как проявление особых функциональных свойств лишь определенных аминокислот [3]. Однако позднее было установлено, что аналогичные, оперантно-детерминирован-ные нейрохимические эффекты способны вызывать и другие вещества органической природы [4-7]. Дальнейший анализ полученных результатов позволил сформулировать в качестве рабочей гипотезы предположение о том, что и в естественных условиях на уровне нейронов интактной ЦНС в период генерации потенциалов действия складываются все необходимые предпосылки для реализации инструментальных форм нейрохимического взаимодействия [8]. В свете этих представлений принципиально важное значение для решения многих актуальных задач, стоящих в области физиологии нервной клетки, приобретает установление максимально полной картины качественно различных процессов, протекающих во время спайковой нейронной активности. Цель настоящей работы - рассмотреть один из аспектов этого круга вопросов, а именно выяснить основные характеристики аминокислотного обмена нервной клетки в период генерации одиночного потенциала действия.
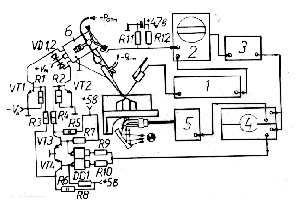
Для решения поставленной задачи проводили авторадиографическое определение интенсивности поглощения отдельным нейроном меченных изотопами аминокислот в условиях локального их подведения в микроинтервалах времени, соответствующих определенной фазе ряда последовательно регистрируемых биоэлектрических разрядов [9]. Объектом изучения были идентифицированные клетки ППа1 и ППа4 полуинтактного препарата ЦНС виноградной улитки Helix Lucorum. При проведении экспериментов применяли быстродействующие микроаппликаторы специальной конструкции [10]. Схема их включения представлена на рис. 1.
Рис.1. Схема коммутации электронных устройств, обеспечивающих работу микроаппликатора. 1 - усилитель биопотенциалов М-707 (WPI); 2 - осциллограф VC-9 (Нихон Кохден);
3 - триггер; 4 - двухканальный электростимулятор 302-Т (WPI); 5 - фотостимулятор (Нихон Кохден); 6 - сельсин микроаппликатора (Н338/6); DD1 - К555 ЛА9; УТ1,2 - КТ 540А;
УТ3,4 - КТ 805; УД1,2 - Д246; R1-R4 - 75 ом; R5,6 - 24 ом;
R7,8 - 510 ом; R9,10 - 1,3 ком; R11,12 - 15 ком.
В основной серии опытов у каждой из исследуемых клеток отводили 1000 потенциалов действия, что соответствовало в среднем 50-минутному периоду ее регистрации. Продолжительность аппликации импульсной изотопной метки во время отдельного спайка составляла 10 мс. В качестве подводимых веществ использовали D,L(2,3H)-лейцин и (2,3Н)-глицин (10 мкКи/мл; 1,2 мкмоль/л), приготовленные на растворе Рингера для холоднокровных (рН = 7,4).
После завершения эксперимента нервную ткань замораживали при Т = -70 оС, а затем на 12 часов помещали в абсолютный этиловый спирт (Т = -20 оС). Срезы толщиной 10 мкм монтировали на предметных стеклах, которые после этого покрывали тонким слоем фотоэмульсии типа Р. Время экспозиции радиоавтографов при Т = +4 оС составляло 10 дней. При проявке использовали проявитель D-19, а в качестве фиксажа - 30% раствор гипосульфита. Интенсивность поглощения изотопов оценивали путем подсчета количества зерен восстановленного серебра над контуром исследуемой клетки. При статистической обработке полученных данных применяли t-критерий Стьюдента, используя компьютерную программу LМикростат¦.
В общей сложности было исследовано 160 нейронов правого париетального ганглия ЦНС виноградной улитки. У 80-ти из них (40 ППа1 и 40 ППа4) анализировалась микровременная динамика трансмембранного переноса лейцина. Результаты исследования 80 других нейронов позволили установить аналогичные характеристики переноса другой аминокислоты - глицина.
Основные полученные данные представлены в таблице.
Первое, что обращает на себя внимание при рассмотрении установленных закономерностей - наличие быстропротекающих изменений интенсивности поглощения указанных аминокислот в период генерации потенциала действия. Процесс этот, как выяснилось, характеризуется устойчивой микровременной динамикой, а именно: в период деполяризации плазматической мембраны наблюдается относительное снижение скорости трансмембранного переноса, после чего имеет место резкое возрастание его интенсивности. Причем величина такого рода изменений для разных аминокислот оказывается существенно различной (р<0,01), что позволяет говорить о специфичности работы систем активного транспорта разных органических соединений в период генерации одиночного потенциала действия. Необходимо особо отметить, что в межспайковые интервалы времени показатели скорости поглощения обеих аминокислот достоверно не различаются (p>0,05). Такого рода отличия являются особенностью именно периода реализации специализированной функции нейрона.
Таблица
Результаты авторадиографического определения интенсивности поглощения меченных изотопами аминокислот в межспайковых интервалах времени и в период генерации потенциала действия. Представлены данные о количестве зерен восстановленного серебра над контуром исследованных нервных клеток. Объем выборки в каждом случае составляет n = 10.
Несомненный интерес представляет и тот факт, что установленные изменения по своей продолжительности значительно превосходят длительность самого потенциала действия. Более того, максимум интенсивности трансмембранного переноса аминокислот приходится на постспайковые интервалы времени. Т.е. наиболее выраженные изменения, связанные с генерацией потенциала действия, происходят в постактивационные (в традиционном понимании) периоды. Эти данные, помимо всего прочего, свидетельствуют о достаточно сложном механизме трансмембранного переноса аминокислот, который не сводится к простому их котранспорту (симпорту) [11] с поступающими в цитозоль нервной клетки ионами Na. В настоящей работе не ставилась специальная задача выяснить конкретные молекулярно-биологические основы реализации этого процесса. Пока можно лишь констатировать, что по времени он совпадает с повышением интенсивности работы Na-K АТФ-азных систем, обеспечивающих восстановление нарушенного в ходе генерации потенциала действия равновесного распределения ионов внутри и вне клетки.
Особого внимания здесь заслуживает то обстоятельство, что для разных по своей специализации нервных клеток конкретные временные показатели указанного процесса оказываются различны. Если для нейрона ППа1 пик интенсивности поглощения обеих аминокислот приходится на 15-ую мс от начала генерации потенциала действия, то аналогичный показатель для нейрона ППа4 уже превышает 30 мс. Кроме того, в последнем случае более продолжительным является и сам период усиления трансмембранного переноса аминокислот.
В свете всех этих данных разрядную активность различных нервных клеток приходится уже рассматривать как последовательность быстропротекающих процессов, идентичных только по основным своим биоэлектрическим характеристикам. Что же касается работы систем активного транспорта органических соединений, то для разных по специализации нейронов их характеристики оказываются существенно различными. Более того, как выяснилось, такого рода отличия (правда в значительно менее выраженной форме) обнаруживаются и у идентифицированных нейронов одного типа в зависимости от средней частоты их разрядов. На рис. 2 представлены результаты соответствующего корреляционного анализа, проведенного для нейронов ППа4, которые имеют достаточно большой разброс средней частоты фоновой импульсации. Установлено, что ее увеличение сопровождается снижением амплитуды постспайкового трансмембранного переноса обеих аминокислот. Т.е. в данном случае проявляется статистически достоверная (p<0,05) обратно пропорциональная корреляционная зависимость между сопоставляемыми рядами параметров. Причем величина коэффициента корреляции для показателей интенсивности поглощения лейцина (R = -0,82) оказывается выше, чем у другой аминокислоты - глицина (R = -0,63).

Рис.2. Зависимость интенсивности
трансмембранного переноса 3Н-лейцина (1) и 3Н-глицина (2) в постспайковые
периоды времени от текущей частоты разрядной активности нервной
клетки. По оси абсцисс - средняя частота генерации потенциалов
действия, имп/сек. По оси ординат - количество зерен восстановленного
серебра над контуром исследуемого нейрона.
В целом, рассмотренные выше данные определенно свидетельствуют о том, что генерация нервной клеткой потенциалов действия, помимо известных мембранно-ионных феноменов, сопровождается возникновением быстропротекающих изменений интенсивности трансмембранного переноса аминокислот. Уже сам по себе этот факт доказывает, что на соматическом уровне нейрона имеются специализированные механизмы, обеспечивающие реализацию экстренной транспортной функции в отношении не только ионных компонентов, но и некоторых органических соединений. Сегодня пока еще трудно судить о роли различных молекулярно-биологических факторов в построении указанной формы клеточной активности, но уже ясно, что, в отличие от ионной составляющей, параметры этого процесса могут меняться в широких пределах даже у сходных по специализации нейронов.
Полученные в настоящей работе результаты, разумеется, отражают лишь некоторые аспекты функциональных свойств Lвозбужденной¦ плазматической мембраны нервной клетки. Но, хотя речь идет только о первых шагах в данном направлении, установленные закономерности уже дают общие представления о характере организации указанных процессов, а также о возможной их роли в развертывании классических и инструментальных форм нейрохимического взаимодействия в ЦНС человека и животных.
Наиболее важным в этом плане заключением можно считать вывод о том, что генерация потенциала действия представляет из себя значительно более сложное во всех отношениях нейрофизиологическое явление, чем простая мембранно-ионная реакция клетки на пороговые изменения уровня ее медиаторного притока. Имеется в виду не только сложность молекулярно-биологической организации трансмембранного переноса. Более широкий подход, прежде всего, предполагается при оценке временных параметров разрядной деятельности нейрона. В свете установленных данных спайковую форму нейронной активности приходится уже рассматривать как процесс, который охватывает значительно большие интервалы времени, чем период собственно биоэлектрических проявлений. Причем весьма вероятно, что этот вывод относится и к завершающим стадиям генерации потенциала действия, и к начальным его этапам, которые связаны с градуальным переходом от Lфонового¦ уровня нейромедиаторного притока к пороговым значениям. Основания для такого предположения дают результаты исследований, свидетельствующие о медиаторно-зависимом характере ряда базовых внутриклеточных механизмов, избирательно чувствительных именно к подпороговым (предпороговым) изменениям концентрации нейромедиаторов в экстраклеточной среде [12]. Последнее обстоятельство, как уже отмечалось [10], имеет принципиально важное значение при рассмотрении различных нейрохимических процессов, развертывающихся в рамках классической (хемореактивной) схемы их построения.
Вместе с тем, установленные закономерности вполне
определенно указывают на необходимость развития более широкой
системы представлений и о механизмах возникновения оперантно-детерминированных
форм вызванной нейронной активности. Как показывают полученные
данные, реализация этого процесса вполне вероятно может осуществляться
не только в ходе взаимодействия нескольких систем вторичных посредников
[7], но и в результате прямого (неопосредованного специализированными
рецепторными образованиями плазматической мембраны) влияния метаболических
факторов на работу электрогенных механизмов нервной клетки.
1. Бобровников Л.В. Исследование корковых нейронов методом микроионофореза, управляемого нейронной активностью // Журн. высш. нервн. деят.- 1986.- ¦ 5.- С.975-977.
2. Бобровников Л.В. Разрядная активность нейрона как фактор реорганизации его нейромедиаторного притока // Мат. VII Междунар. конф. LРазвитие общей теории функциональных систем¦.- М., 1986.- С.7-9.
3. Bobrovnikov L.V. Microiontophoretic applycation of L-glu-tamate as a factor of operant conditioning of neuron firing pattern // Systems Research in Physiology.- 1991.- V.4.- P.219-228. Gordon and Breach Science Publishers. New York.
4. Belluzzi J.D., Kossuth S.R., Lam D., Derakhshanfar F., Shin A., Stein L. Cocaine self-administration patterns: dupliсаtion by combination of dopamine D1 and D2 agonists // Society for Neuroscience Abstracts.- 1993.- ¦ 19.- P.1862.
5. Stein L., Xue B.G., Belluzzi J.D. A cellular analogue of operant conditioning // Journal of the Experimental Analysis of Behavior.- 1993.- ¦ 60.- P.41-53.
6. Xue B.G., Belluzzi J.D., Stein L. In vitro reinforcement of hippocampal bursting activity by the cannabinoid receptor agonist CP-55,940 // Brain Research.- 1993.- V.626.- P.272-277.
7. Stein L., Xue B.G., Belluzzi J.D. In vitro reinforcement of hippocampal bursting: a search for Skinner^s atoms of behavior // Journal of the experimental analysis of behavior.- 1994.- ¦ 61.- P.155-168.
8. Бобровников Л.В. Репарационная активность и нейрогенез // Журн. высш. нервн. деят.- 1996.- ¦ 5.- С.961-964.
9. Бобровников Л.В. Способ определения проницаемости мембраны нейрона // Патент на изобретение ¦ 1790768 G 01 N 33/48 1992 г. (Дата приор. 16.09.89 г.).
10. Бобровников Л.В. LПодпороговые¦ нейромедиаторные процессы и их роль в реализации механизмов интегративной деятельности нейрона // Вестник новых медицинских технологий.- 1998.- ¦ 1. - в печати.
11. Erecinska M. The neurotransmitter amino acid transport systems // Biochem. Pharmacol.- 1987.- V.36, ¦ 21.- P.3547-3555.
12. Коштоянц Х.С. Энзимохимическая гипотеза
возбуждения // Физиологич. журн. СССР.- 1950.- ¦ 1.- С.92-96.
It has been found, that the process of action potential generation include the fast alteration of the amino acid uptake. On the base of this date the role of the non-neurotransmitter substances in the neuronal activity formation is discussed.