
УДК 612.821.6; 615.78
Регуляторные пептиды (РП) - это важный класс эндогенных соединений. Они имеют большое значение в формировании различных физиологических и эмоциональных состояний организма: жажды, голода, боли, агрессии, страха и др. Имеются экспериментальные данные [1, 2, 3] о том, что многие из эндогенных регуляторных пептидов существуют в организме не только в свободной форме, но и в виде комплексов ковалентно или химически связанных с белками крови и других биологических жидкостей носителями (вазопрессин, окситоцин, гастрин, холецистокинин и др.).
Оценивая функции РП, авторы, как правило, имеют в виду активность свободных форм биорегуляторов данного типа. Вместе с тем, рассматривая значение эндогенных конъюгатов РП с белками, указывают лишь на транспортную и т. н. Lде-понирующую¦ функции этих соединений, обеспечивающих защиту РП от быстрого протеолиза и выведения из организма.
Однако имеются основания считать, что физиологическая роль эндогенных конъюгатов РП с белками этим не ограничивается. Можно предположить, что такого рода соединения имеют самостоятельные функции в молекулярных процессах регуляции гомеостаза, формирования и реализации целенаправленного поведения различной биологической модальности.
В связи с этим представляется несомненно актуальным исследование самостоятельной физиологической роли конъюгатов РП с белками крови и внутриклеточными белками (как аналогов эндогенных конъюгатов) в процессах системной организации физиологических функций.
Основной целью настоящей работы явилось изучение
участия аналогов эндогенных, конъюгированных с белками (сывороточный
альбумин, катионизированный сывороточный альбумин) форм ангиотензина-II
(A-II) в реализации целенаправленного поведения.
В первой серии экспериментов (36 крыс популяции Wistar, по 12 животных в каждой группе) был проведен сравнительный анализ физиологической активности нативного A-II и конъюгатов A-II с белками-носителями - бычьим сывороточным альбумином (БСА) и катионизированным БСА. У всех животных динамически оценивали суточное количество потребляемой пищи и воды, массу тела, двигательную активность и артериальное давление (АД) до и после периферического (подкожного - п/к и внутриартериального - в/а) введения A-II и конъюгатов. A-II вводили в дозах 0,5, 1, 5, 10, 20, 50 и 100 мкг. Конъюгаты A-II вводили в дозах, содержащих эквивалентные количества A-II, химически связанные с носителями. Контролем служило введение 0,5 мл физиологического раствора.
АД регистрировали как прямым методом в хвостовой артерии (12 животных), так и непрямым манжеточным способом.
Синтез конъюгата A-II с БСА был осуществлен М.Ф.Обуховой с использованием карбодиимидного метода. Полученный таким методом конъюгат содержал 10-12 молекул пептида на 1 моль белка. Конъюгат A-II с катионизированным БСА (7 молей A-II на 1 моль катионизированного БСА, pI = 9,0) был предоставлен О.А.Гомазковым (Институт биомедицинской химии РАМН). Катионизированный БСА, в отличие от немодифицированного БСА, является белком-носителем, придающим всей молекуле конъюгата в целом электростатические свойства. В опытах использовали A-II производства фирмы Serva.
Эксперименты второй серии были выполнены на 18 крысах-самцах (12 опытных и 6 контрольных) популяции Wistar, предварительно обученных выполнению Lманипуляторных¦ поведенческих актов для получения воды с использованием разработанного в лаборатории мотиваций НИИ нормальной физиологии им. П.К.Анохина РАМН автоматизированного комплекса технических средств [4], позволяющего объективно оценивать у крыс выполнение ряда последовательных этапов результативного поведения. (рис.1).
По условиям опыта предварительно обученные воднодепривированные животные после помещения в стартовый бокс должны были выполнить побежку в Lманипуляторный¦ бокс и получить в нем порцию воды (0,5 мл). Крысы путем Lманипулирования¦ передними лапами должны были повернуть диск на определенный угол, что обеспечивало автоматическую подачу порции воды в поилку. По окончании приема воды через фиксированный интервал времени передвижная шторка прекращала доступ к вращающемуся диску, включался тредбан и животные должны были возвратиться в стартовый бокс, чтобы иметь возможность вновь повторить целостный поведенческий акт.
LМанипуляторное¦ поведение животных регистрировали с использованием компьютера LAppricot-Xi¦ в виде отдельных файлов на магнитных дисках 3.5" 720 кВ во время 8-минутных сеансов в светлое время суток. После сеансов тестирования крысам предоставляли в течение 15 минут свободный доступ к воде, определяя количество воды, потребляемое каждым животным. В промежутках между сеансами крысы воды не получали и имели свободный доступ к пище.
У животных в установке определяли следующие временные показатели реализации сложных Lманипуляторных¦ поведенческих актов: dT1 - латентный период акта до начала выхода на тредбан; dT2 - период с момента выхода на тредбан до принятия позы Lманипуляции¦; dT3 - время с момента принятия позы - Lманипуляции¦ до начала потребления воды; dT4 - время потребления воды; dT5 - время с момента прекращения потребления воды до выхода из позы манипуляции; dT6 - время возвращения на вход в тредбан; dT7 - время перехода с тредбана в стартовый бокс. Общую длительность каждого акта (dT8) определяла сумма вышеперечисленных показателей.
Статистической обработке по количеству и внутренней структуре подвергались только результативные акты. В ходе статистической обработки вычисляли средние значения временных показателей по каждому дню эксперимента для каждой крысы отдельно. При оценке показателей внутренней структуры поведенческого акта (dT1 - dT7) вычисляли их сравнительную величину в процентах от общей длительности каждого отдельного результативного акта.
При проведении экспериментов были выделены следующие периоды: 1 - фоновое наблюдение; 2 - введение конъюгата А-II с БСА и нативного А-II; 3 - острое введение этанола.
В ходе проведения экспериментов были выделены следующие группы животных: I группа (6 крыс), которым во 2 периоде осуществляли инъекцию конъюгата А-II c БСА (однократно, п/к, в дозе 150 мкг/кг А-II, химически связанного с белком). II группе (6 животных) вводили однократно, п/к А-II в дозе 150 мкг/кг. Животным III группы аналогичным образом делали инъекции физиологического раствора.
В 3 периоде животным всех групп п/к вводили 10% раствор этанола в дозе 0,5 г/кг массы тела.
Пороги болевой чувствительности определяли после
тестирования в установке через 20 и 40 минут, а также через 1
сутки после инъекции веществ, используя Ltail-flick test¦.
В первой серии экспериментов было обнаружено, что в/а введение конъюгата А-II с катионизированным БСА вызывает отчетливое повышение АД и снижение частоты сердечных сокращений, начиная с доз 0,5 - 1 мкг. Длительность данного эффекта колебалась у разных животных от 7 минут до 1 часа, составляя в среднем 30 мин. (рис. 2 А). У части крыс вслед за гипертензивной фазой было отмечено и гипотензивное действие в течение 3-4 часов. В/а введение нативного А-II приводило к появлению менее выраженного гипертензивного эффекта, длительность которого не превышала 5 минут, и тахикардии (рис. 2 Б). Следует отметить, что последовательное увеличение доз как конъюгата А-II с катионизированным БСА, так и А-II (от 5 до 100 мкг) не вызывало достоверного усиления гипертензии. Наряду с гемодинамическими эффектами введение даже минимальных доз конъюгата А-II с катионизированным БСА (0,5-5 мкг) инициировало потребление воды, пищи, сопровождавшееся снижением двигательной активности, интенсивной моторной стереотипией и грумингом. В то же время при введении нативного A-II наблюдалось умеренно выраженное питьевое поведение, причем только при использовании максимальной дозы (100 мкг).
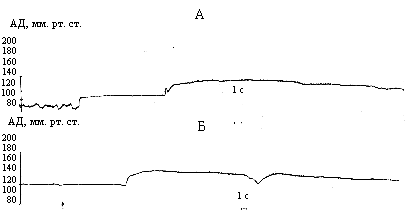
Необходимо подчеркнуть, что гемодинамические и дипсогенные реакции А-II хорошо изучены как при внутримозговом, так и при периферическом введении [5, 6]. В связи с этим можно предположить, что выявленные в наших экспериментах эффекты конъюгата А-II с катионизированным БСА опосредуются не только взаимодействием с периферическими A-II-рецепторами, но и за счет влияния на внутримозговую ренин-ангиотензиновую систему. Это согласуется с данными [7] о том, что пептиды, конъюгированные с катионизированными белками, характеризуются повышенной проницаемостью через гемато-энцефа-лический барьер.
При п/к введении конъюгата А-II с БСА, т.е. с немодифицированным белком-носителем, у крыс наблюдаются аналогичные поведенческие и гемодинамические эффекты, которые, однако, проявляются при использовании значительно более высоких доз (в 40-50 раз). Минимальная эффективная доза конъюгата А-II с БСА по сравнению с нативным А-II в данном случае была значительно меньше, чем минимальная эффективная доза А-II (50 мкг/кг и 300 мкг/кг соответственно).
Более широкий спектр физиологической активности конъюгированных форм A-II по сравнению с нативным A-II обусловлен, по-видимому, различным характером их взаимодействия с рецепторами периферических региональных ренин-ангиотен-зиновых систем [8, 9].
При изучении питьевого Lманипуляторного¦ поведения крыс во второй серии экспериментов было обнаружено, что по общему количеству поведенческих актов в 1 и 2 периодах все группы достоверно не различались. В условиях активации эндогенных опиоидных систем путем острого введения этанола в 3 периоде у животных III группы регистрировали увеличение (р<0,05) количества манипуляторных поведенческих актов за сеанс тестирования. Аналогичное, но менее выраженное увеличение числа поведенческих актов наблюдалось у животных II группы. Только у животных I группы введение этанола практически не изменяло характер реализации сложного питьевого поведения.
Существенными характеристиками для любого целенаправленного поведения являются параметры достигаемого полезного приспособительного результата. В случае изучаемого питьевого поведения таким параметром является объем полученной животным жидкости. В связи с этим определяли объемы воды, выпиваемой каждой отдельной крысой при реализации сформированного навыка в установке, при допаивании в условиях свободного доступа, что является врожденной формой питьевого поведения, а также общий объем потребленной жидкости. Было показано, что у животных всех групп выраженно снижается потребление воды при допаивании в 3 периоде эксперимента. Этот эффект наиболее отчетливо проявляется у крыс I группы.
При изучении тонкой структуры поведения оказалось, что длительность реализации центрального этапа поведенческого акта, в наибольшей степени связанного с удовлетворением базовой для данной экспериментальной модели питьевой мотивации, - время потребления порции воды (dT4), в наибольшей степени связанное с нейрофизиологическими процессами подкрепления, - в условиях активации эндогенных опиоидных систем путем острого введения этанола значительно возрастала лишь у крыс III группы (р<0,05), не изменяясь существенно у животных других групп (рис. 3 А). Аналогичные изменения показателя dT4, были выявлены в наших предыдущих исследованиях в условиях хронического введения морфина [10].
В то же время у животных II и III групп на протяжении эксперимента
последовательно уменьшалась длительность показателя dT7, характеризующего
заключительный этап поведенческого акта, а у крыс I группы данный показатель изменялся
противоположным образом (рис. 3 Б).
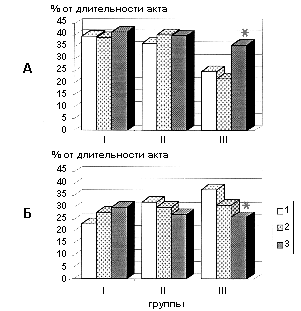
Таким образом, периферическое введение конъюгата А-II с БСА отчетливо подавляет процесс автоматизации предварительно выработанного Lманипуляторного¦ поведенческого акта. Это проявлялось в том, что на протяжении всего периода наблюдения у животных не возрастало количество результативных поведенческих актов, последовательно увеличивалась длительность выполнения заключительного этапа поведенческого акта.
В условиях активации эндогенных опиоидных систем путем острого введения этанола инъекция конъюгата А-II с БСА не изменяла сложившийся характер реализации сложного питьевого поведения у крыс. При этом реализация врожденной формы питьевого поведения подавлялась в наибольшей степени по сравнению с животными, которым вводили А-II и физиологический раствор.
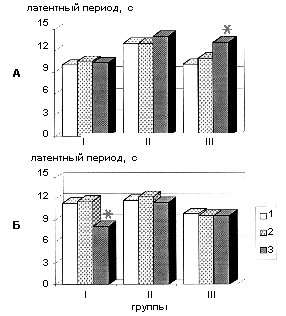
Определение порогов болевой чувствительности показало, что в условиях активации эндогенных опиоидных систем путем острого введения этанола введение конъюгата А-II с БСА и А-II (I и II группы крыс) не вызывало изменений порогов болевой чувствительности в отличие от крыс III группы, у которых через 40 минут после введения этанола было отмечено достоверное (p<0,05) снижение болевой чувствительности (рис. 4 А). Было обнаружено, что только у животных, которым вводили конъюгат А-II через 1 сутки после введения этанола существенно снижался латентный период отдергивания хвоста (р<0,05), что свидетельствует о развитии гипералгезии (рис. 4 Б) и обусловлено, по-видимому, длительным действием конъюгата А-II с БСА, т.к. конъюгированные формы пептидов являются устойчивыми к действию эндопептидаз [11], и их взаимодействие с рецепторами нативного пептида существенно продлевается и носит иной характер.
Таким образом, полученные данные указывают на наличие тесных функциональных взаимоотношений эндогенных ренин-ангиотензиновой и опиоидных систем организма, которые играют, по-видимому, важную роль в регуляции адаптационных процессов.
Конъюгированные формы А-II имеют, по-видимому, самостоятельные, преимущественно интегративные функции по сравнению с нативным пептидом в системной организации поведения.
Конъюгированные формы А-II характеризуются значительно более широким спектром физиологической активности и пролонгированным действием по сравнению с нативным пептидом, различным образом влияют на врожденные и приобретенные формы питьевого поведения, оказывают модулирующее влияние на процессы подкрепления.
Таким образом, полученные результаты позволяют считать конъюгированные формы пептидов принципиально новым классом эндогенных биорегуляторов, дальнейшее изучение которых открывает новые перспективы в исследовании механизмов действия физиологически активных веществ, процессов регуляции гомеостаза в организме.
Представляется, что на основе аналогов таких конъюгатов
возможно создание новых типов лекарственных средств.
An a wide spectrum in behavioral activity of conjugates of angiotensin-II with bovine serum albumin (A-II BSA) was demonstrated. Feeding behavior, intensive motor stereotypy, prolonged hypertension after A-II BSA administration were obtained as a specific dipsogenic effects (both A-II BSA ad native A-II).
Automatization of learning forms in drinking behavior decreased under A-II BSA peripheral injections. Natural forms of drinking behavior were suppressed while a hyperalgesic effects after ethanol administration registered.
The real integrative functions of natural A-II-protein
complexes in organism were suggested (comparing with native A-II).
Андреев Дмитрий Анатольевич,
1976 года рождения. Студент 4-го курса Московской медицинской
академии им. И.М.Сеченова. Активно работает в студенческом научном
кружке кафедры нормальной физиологии ММА им. И.М.Сеченова. В течение
последних трех лет проводит самостоятельные экспериментальные
исследования в лаборатории мотиваций НИИ нормальной физиологии
им. П.К.Анохина. Основная область научных интересов: разработка
объективных методов оценки поведения животных, нейроиммунология.
Певцова Елена Игоревна.
Окончила в 1976 г. Московский Государственный Университет им.
М.В.Ломоносова. После обучения в аспирантуре на кафедре физиологии
человека и животных биологического факультета МГУ и защиты диссертации
работает в НИИ нормальной физиологии им. П.К.Анохина. Старший
научный сотрудник лаборатории мотиваций. В настоящее время занимается
изучением пептидергических и иммунологических аспектов формирования
и реализации целенаправленной поведенческой деятельности животных.
Толпыго Светлана Михайловна.
В 1977 г. окончила 1 Московский медицинский институт им. И.М.Сеченова.
Кандидат медицинских наук, ведущий научный сотрудник лаборатории
мотиваций НИИ нормальной физиологии им. П.К.Анохина РАМН. Автор
более 40 научных работ. Основные направления научной деятельности:
изучение нейрофизиологических и нейрохимических механизмов различных
форм биологических влечений, физиологического действия регуляторных
пептидов, разработка иммунологических подходов анализа целенаправленного
поведения животных.