УДК 612.821.2+612.822+615.37
В настоящее время разработаны новые подходы к изучению процессов неврологической памяти [1, 2]. Одним из них является применение классических и новых (вновь синтезированных или выделенных) иммуномодуляторов для улучшения процессов запоминания и воспроизведения из памяти [3, 4]. Эти подходы основываются на многочисленных исследованиях [5], позволяющих говорить об общности происхождения, функционирования и взаимосвязях нервной и иммунной систем организма. С другой стороны, развитие молекулярно-генетических методов исследований приводит к несомненному заключению о единстве принципов организации клеточных взаимодействий во всех системах организма, основанных на законах комплементарных отношений [6, 7].
Эти данные приводят к необходимости изучения и анализа системных механизмов действия иммуномодуляторов на процессы центральной регуляции жизнедеятельности организма.
В предыдущих исследованиях лаборатории было показано, что в структуре паттерна импульсной активности нейронов находят отражение доминирующие мотивационные состояния и подкрепляющие воздействия. Кроме того, были обнаружены нейрофизиологические корреляты, отражающие стадии формирования функциональной системы на моделях обучения пищедобывательному, питьевому и оборонительному поведению [8].
Исходя из вышеизложенного, цель настоящего исследования заключалась в изучении воздействия ряда иммуномодуляторов (фрагмента 2-интерферона RITLY, интерлейкина-1b и нейротропина - НТ) в процессе выработки оборонительного навыка у кроликов и крыс с анализом поведенческих, вегетативных и нейронных показателей.
Первая серия экспериментов проведена на 42 крысах-самцах линии Вистар: 21 контрольных и 21 опытных. Опытная группа была подразделена на три подгруппы: а) с внутрижелудочковым введением 0,1 мкг фрагмента 125-129 2-интерферона RITLY в 2 мкл дистиллированной воды (n = 6); б) с внутрижелудочковым введением интерлейкина-1b в дозе 100 нг в 10 мкл физиологического раствора (активность 3107 ЕД/1 мкл) (n = 8); в) с интраперитонеальным введением 0,2 мл официнального раствора нейротропина.
Пентапептид RITLY расположен в участке петли 122-144, соединяющей два -спиральных домена молекулы 2-интерферона, и является одним из ее активных центров. Кроме того, данный пентапептид относится к так называемому Lконсервативному¦ участку, встречающемуся без изменения последовательности аминокислот у всех молекул многочисленного семейства -интерферонов. Структурный и функциональный анализ показал, что RITLY имеет определенное сходство с тимопентином и спленопентином, высокоактивными фрагментами тимопоэтина и спленина и обладает иммуномодулирующим эффектом [9].
Нейротропин (NSP) является иммуномодулятором эндогенного происхождения небелковой природы и представляет собой коньюгированную смесь олигосахаридов, выделенных из кожи кроликов при развитии гиперчувствительности замедленного типа, вызванной вирусом оспы (Nippon Zoki, Япония). Нейротропин обладает широким спектром действия, максимально проявляющимся при необходимости компенсации нарушенных функций. Отмечают его иммуномодулирующее, антиаллергическое, антистрессорное, анальгетическое действия [10]. Центральное действие нейротропина проявляется в усилении 2-ритма ЭЭГ, изменении активности нейронов гипоталамических ядер, сенсомоторной коры, гиппокампа [11].
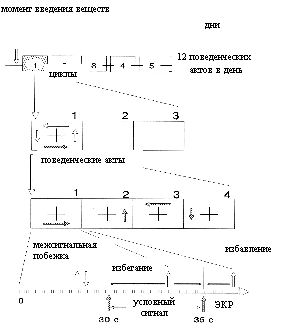
Обучение крыс активно оборонительному поведению проводилось с помощью автоматизированного комплекса, включающего 4-камерный лабиринт на базе OPTO-VARIMEX и ЭВМ LApple-II¦, осуществляющего управление и обработку сигналов. Схема проведения эксперимента показана на рис. 1. Изучаемые вещества и физиологический раствор или дистиллированная вода, в соответствующих объемах и способах введения (в контрольных группах), вводились непосредственно перед первым сеансом обучения.
Регистрировались параметры результативности (количества избеганий, избавлений, ошибок) и межсигнальной активности крыс.
Во второй серии экспериментов исследовалась динамика изменений вегетативных (ЭКГ и дыхание) показателей и активности нейронов поля СА1 гиппокампа у 27 кроликов породы шиншилла в процессе выработки оборонительного поведения.
Обучение состояло из 30 сочетаний условного сигнала (4 звуковых щелчка, через 1 с) и электрокожного раздражения задней лапы животного силой тока 20 млА, длительностью 70 мс, совпадающим с 4-м щелчком. Сочетания предъявлялись в случайном режиме времени через 40-90 с. За 3-5 мин. до начала обучения вводились: а) дистиллированная вода (10 мкл) в боковой желудочек головного мозга (контроль-1, n = 5) или физиологический раствор внутривенно 0,5 мл (контроль-2, n = 10); б) RITLY в боковой желудочек головного мозга в дозе 1 мкг в 10 мкл дистиллированной воды (n = 5); в) 0.5 мл официнального раствора нейротропина внутривенно.
Обработку нейронной активности и кардиоинтервалограмм
проводили по специально разработанным нами программам, включающим
расчет среднестатистических показателей, гистограмм распределения
и актокорреляционных функций последовательности межимпульсных
или RR-интервалов. Сравнение межгрупповых различий осуществлялось
с помощью критерия t-Стьюдента и факторного анализа.
Проведенные исследования показали, что по мере обучения активно-оборонительному навыку результативность у крыс, в частности количество избеганий, возрастает во всех группах (рис. 2). Однако достоверно (p<0,05) более высокие значения по сравнению с контрольными группами наблюдались на 2-й и 3-й день обучения при введении RITLY и на 3-й день при введении нейротропина по критерию t-Стьюдента и за весь период обучения при факторном анализе. При введении интерлейкина-1b обнаружено достоверное (p<0,05) снижение количества избеганий на 2-й и 6-й дни обучения.
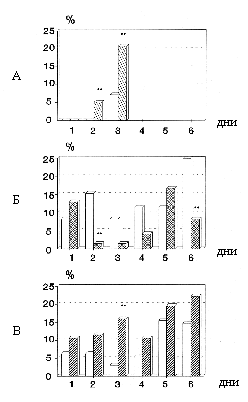
Более высокой результативности животных опытных групп соответствовала и низкая ошибочность. При этом, как показал анализ, крысы совершали достоверно (p<0,05) меньшее число ошибок, связанных с неправильным выбором направления к безопасному отсеку.
В процессе обучения у всех крыс наблюдались межсигнальные побежки (МСП) в неосвещенные отсеки с электрокожным подкреплением до подачи условного сигнала. Среднее количество МСП за весь период обучения достоверно не различалось по группам. Анализ динамики взаимосвязи межсигнальной активности с последующей результативностью в этих циклах позволил обнаружить следующие факты. В контрольной группе животных (рис. 3А) в первые три дня обучения МСП равновероятно встречались как в ошибочных, так и в результативных поведенческих актах. Однако с 4-го дня происходило возрастание числа МСП и достоверно (p<0,05) более высокая их корреляция с правильным выполнением оборонительного акта.
В двух опытных группах, с введенным RITLY
(рис. 3Б) и нейротропином (рис. 3Г), достоверная положительная
взаимосвязь (p<0,05) МСП с результативностью наблюдалась
с первого же дня обучения и сохранялась на всем периоде наблюдений.
Таким образом, введение животным иммуномодуляторов фрагмента 2-интерферона RITLY и нейротропина способствовало обучению активно-оборонительному поведению и одним из механизмов их действия являлась активация процессов межсигнального воспроизведения из памяти вырабатываемого навыка.
Анализ нейрофизиологических показателей при формировании оборонительного поведения у кроликов позволил выявить аналогичное влияние иммуномодуляторов RITLY и нейротропина на процессы обучения.
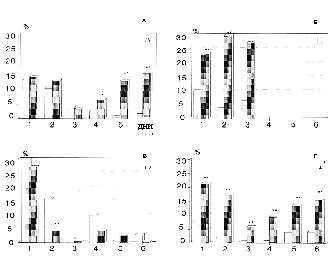
Было обнаружено, что изменения ритма дыхания и кардиоинтервальных характеристик на условный сигнал возникали достоверно (p<0,001) раньше (5-7 сочетания) в опытных группах (рис. 5А) , по сравнению с контрольными животными (12-15 сочетания) (рис. 4А).
В этой же серии экспериментов была зарегистрирована активность 83 нейронов поля СА1 гиппокампа: 38 - в контрольной группе (12 кроликов), 18 - при введении RITLY (5 кроликов) и 27 - при введении нейротропина (7 кроликов).
Анализ структуры импульсной активности нейронов,
в которой выделялись тормозная, нерегулярно-пачечная и регулярная,
во время условного стимула за весь период обучения позволил выявить
различия между подопытными группами и контролем. Оно заключалось
в том, что по сравнению с нейронами контрольной группы, где преобладали
ответы с нерегулярно-пачечным типом активности (57,2%), в экспериментальных
группах этот тип был представлен лишь в 28% нейронов при введении
RITLY и 11% на фоне нейротропина. Вместе с тем, в экспериментальных
группах преобладали нейроны с тормозной (32%-RITLY и 48,2%-НТ)
и регулярной (40%-RITLY и 40,7%-НТ) формой активности по
сравнению с контролем (23% - торможение и 15% - регуляризация).
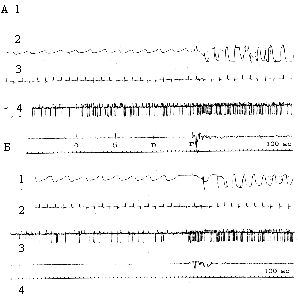
Изменения этих же параметров при межсигнальной активности (Б). а, б, в - первые три условные звуковые сигналы;
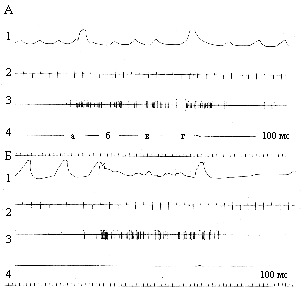
Рис. 5. Изменения
нейрофизиологических показателей при
обучении животного в условиях действия нейротропина
(А) и при межсигнальной активности (Б). Обозначения - как на
рис. 4 Регулярная
и тормозная формы активности нейронов характерны для подкрепления
и по мере обучения возникают на 7-10 и 25-30 условный сигнал в
контроле, соответственно. В экспериментальных группах эти формы
активности наблюдались достоверно (p<0,05) на более
ранних стадиях обучения по сравнению с контролем.
Помимо исследования вегетативных и нейронных показателей
на влияние условного сигнала и электрокожного болевого подкрепления
нами были проанализированы межсигнальные периоды. Было обнаружено,
что у животных всех групп при обучении возникала кратковременная
межсигнальная активность (МСА), которая выражалась в резком изменении
дыхания, кардиоритма и нейронной активности, а также сопровождалась
движениями животного. МСА в опытных группах характеризовалась
повторением тех временных и структурных параметров кардиоритма,
дыхания и импульсной активности нейронов, которые были характерны
для сочетаний условных и подкрепляющих воздействий. Примеры МСА
в контрольной и экспериментальных группах представлены на рис.
4Б и рис. 5Б.
Межсигнальная активность, с точки зрения теории
функциональных систем, представляет собой опережающую поведенческую
деятельность, связанную с процессами воспроизведения из памяти
тех исполнительных механизмов, которые приводили к положительному
результату. Она отражает срыв Lпредпусковой интеграции¦ на стадии
афферентного синтеза и воспроизводит эфферентную программу действия
.
Таким образом, можно предположить, что в условиях
действия иммуномодуляторов при обучении животных активируются
процессы воспроизведения навыка из памяти, которые проявляются
в межсигнальных изменениях деятельности центральных (нейрон),
периферических (дыхание, сердечная деятельность) и двигательных
компонентов, обеспечивающих результативность в функциональной
системе оборонительного поведения.
На основании полученных нами данных о влиянии ряда
иммуномодуляторов на процессы обучения необходимо также указать
на некоторые их различия, которые вероятно обусловлены нейрохимическими
особенностями их участия в этих процессах. Это находит подтверждение
в литературных данных о том, что действие веществ ряда интерферонов
и нейротропина опосредуется их взаимодействием с опиатной системой
головного мозга, которая участвует в механизмах подкрепления [12].
Тогда как показано, что интерлейкины играют определенную роль
в механизмах развития стресса за счет модуляции уровня гипоталамического
гистамина и активации простагландина Е-2 [13, 14]. В некоторых
других моделях обучения животных оборонительному поведению, в
которых обстановочная афферентация менее стрессогенна, интерлейкин-1b
может оказывать положительное влияние (Журавлев Б.В., Муртазина
Е.П. и Сулин В.Ю., в печати).
Immunomodulators were used as a method of influence
on the processes of memory and retention in the experimental model
of learning to the active-avoidance behavior. It was shown that
administration of fragment of -2 interferon RITLY and neurotropin
relieved the learning of rats and rabbits. Intertrial activity
observed during the learning has significant higher levels in
the experimental groups and was accompanied by changes of neuronal
activity, respiration, variation of heart rate and motor activity.
These changes were similar to those during the conditional and
unconditional signals. It was found that success of learning
correlated with the levels of intertrial activity. These results
suggest that immunomodulators activate the retrival processes
and lead to more steady consolidation of avoidance behavior.
Журавлев Борис Васильевич,
1943 года рождения. В 1967 году окончил 1 Московский медицинский
институт им. И.М.Сеченова. Доктор медицинских наук, профессор.
Работает руководителем лаборатории общей физиологии функциональных
систем НИИ нормальной физиологии им.П.К.Анохина РАМН. Автор более
100 научных работ. Является специалистом в области системной нейрофизиологии
поведения, мотиваций, памяти и эмоций. В последнее время разрабатывает
проблему нейроиммунологических процессов запоминания и воспроизведения
из памяти.
Муртазина Елена Павловна,
1962 года рождения,- старший научный сотрудник НИИ нормальной
физиологии им.П.К.Анохина РАМН. В 1987 году закончила медико-биологический
факультет 2-го МОЛГМИ им. Н.И.Пирогова. В 1993 по окончании аспирантуры
защитила кандидатскую диссертацию. Научные интересы - системная
нейрофизиология поведения животных и человека. Имеет 15 публикаций.
Сулин Валерий Юрьевич,
1962 года рождения,- доцент кафедры физиологии человека и животных
Воронежского Государственного Университета. Закончил аспирантуру
при НИИ нормальной физиологии им. П.К.Анохина РАМН в 1993 году.
Кандидат биологических наук. Научные интересы лежат в области
изучения проблем нейрофизиологии и психофизиологии поведения человека
и животных.